



Популяция пшеницы, как и других растений, среди прочих характеристик обладает стабильностью развития, которая определяется по величине флуктуирующей асимметрии (ФА) листовой пластины, как разности в величине левого и правого билатерально-симметричных признаков.
Ранние работы в области стабильности развития и асимметрии проводились с использованием линейных или счетных признаков или сравнением площади половинок листа. При таком подходе обозначалась ось симметрии, строились перпендикулярные отрезки. Величина флуктуирующей асимметрии определялась по абсолютной разности, отнесенной к сумме значений признаков. Из-за продольного строения пластины злаковых, требуется достаточно большое количество измеряемых отрезков, а дальнейшие статистические процедуры включают проверку на нормальность распределения и статистическую оценку результатов. В последнее время созданы программы для автоматического сканирования и расчета асимметрии различных листьев, в том числе злаковых культур [1]. Морфологические характеристики, такие как форма, остаются за пределами такого подхода, который носит скорее описательный, но не аналитический характер.
Авторами был выбран метод геометрической морфометрии, который удобен тем, что позволяет сравнить асимметрию по форме края правой и левой половины листа. Согласно этому методу, метки с координатами ХУ расставляются равномерно по контуру изображения листовой пластины, способствуя увеличению числа степеней свободы df. Программа MorphoJ [2] регистрирует как общую изменчивость координат меток (компонент симмтерии), так и билатерально-симметричную изменчивость координат парных меток (компонент асимметрии).
Исследование ФА в листьях древесных растений показало высокое содержание направленной асимметрии (НА), которое, как правило, искусственно увеличивает флуктуирующую асимметрию. В лесных ценопопуляциях ФА, как нормальная изменчивость листовых пластин, варьирует в широком диапазоне из-за неучтенных факторов, среди которых наиболее важный – освещение. Фактор урбанизации согласно недавним сообщениям оказывает серьезное влияние на снижение стабильности развития в популяциях березы повислой [3, 4].
Флуктуирующая асимметрия зависит от различных причин. Привлекают внимание генетические и эпигенетические факторы, хотя основное влияние оказывает среда, а саму ФА относят к фенотипической изменчивости, отдельно выделяя флуктуационную изменчивость [5]. В предыдущих работах была проведена оптимизация числа расставляемых меток и объема выборки [6, 7].
Длина листовых пластин заслуживает отдельного рассмотрения, так как изменчивость длины пластины ведет к возможному изменению как формы, так и асимметрии. Известно, что продолжительность вегетации озимых злаков длиннее, чем у яровых форм, отсюда ожидаемо большие размеры пластин у озимой пшеницы и большая величина ФА, хотя линейная характеристика не всегда коррелирует с величиной ФА. Ранее исследования по сравнению стабильности развития пшеницы не проводились или выполнены недостаточно. Например, работы, проведенные О.А. Устюжаниновой с соавт., были направлены на определение коэффициента флуктуирующей асимметрии озимой и яровой пшеницы в зависимости от насыщенности севооборота бобовыми растениями на фоне внесения минеральных удобрений [8, 9]. Таким образом, рассматривалось не менее двух факторов, однако взаимодействие их, как и влияние года посева или метрические характеристики листовой пластины, не учитывались. Не учитывалось также возможное присутствие направленной асимметрии, как частого спутника ФА.
Пшеница, как и другие культурные сорта злаковых – удобные объекты для исследования, из-за равномерного освещения посевов и относительной генетической чистоты семенного материала.
В цель настоящей работы входило:
а) изучение влияния дозы удобрения на длину листовых пластин, их форму и величину ФА;
б) сравнение ФА и направленной асимметрии (НА) в листьях двух сортов пшеницы.
Морфогеометрический (Прокрустов) дисперсионный анализ проводился с построением усредненной конфигурации (центроида) и расчетом дисперсии парных меток по обеим сторонам пластины в сравнении их для определения величины флуктуирующей асимметрии.
Материалы и методы исследования
Флаговые листовые пластины собирались во второй половине июля 2021 г. на территории Федерального аграрного научного центра (г. Суздаль, Владимирская обл.) с координатами 56,2157°N; 40,5166°E. Работа проводилась в рамках многолетнего адаптивно-ландшафтного эксперимента, проводимого на серых лесных почвах Суздальского Ополья с принятой системой севооборота с использованием пшеницы (Triticum aestivum L.1753). Использовалось два сорта: сорт Грань (озимая) и сорт Поэма (яровая).
Интенсивная доза (1-я) имела следующий состав: N60P60K60 для яровой и N60 + 40 т навоза/га + для озимой пшеницы. Высокоинтенсивная (2-я): N90P90K90 для яровой и N60 + 40 т навоза/га + N40P40K40 для озимой пшеницы (индексы указывают на объем в килограммах вещества, вносимого на 1 га).

Рис. 1. Расположение основных (№ 1 и 2) и вспомогательных меток (№ 3–102)
Площадь делянки составляла 35 м2 (5×7), а контролем служили полосы, разделявшие делянки с 1-й и 2-й дозами удобрения и полосы по краю поля. Обе дозы вносились непосредственно перед посевом.
Листовые пластины собирались рандомно, избегая края делянки 1 м шириной. Выбирались сформированные симметричные пластины без изгибов и поражения микроорганизмами с длиной 16–18 см. С каждой делянки (всего четыре) собиралось 25–30 пластин, затем они вымачивались сутки в бытовом 5 %-м детергенте, высушивались и наклеивались на бумагу абаксиальной стороной наверх и сканировались сканером Canon (300dpi) вместе с линейкой, градуированной по ГОСТ. Изображения сохранялись в формате JPG.
Нанесение первой и второй меток (пакет TPSDig2) проводилось в основании пластины, лишенной лигулы (язычка), и в верхушке пластины (апекс). Вспомогательные метки наносились по 50 по обоим краям – левому и правому. Эти метки считались полуметками, в отличие от первых – истинных меток, образующих ось симметрии (рис. 1).
Построение кривых проводилось первоначально нанесением 25–30 меток (outlines). Затем их число доводилось до 50 и проводилось выравнивание расстояний между ними. Такая процедура проводилась четыре раза: два раза по часовой и два раза – против часовой стрелки.
После определения координат всех меток и создания общего файла проводилась выбраковка выбросов, т.е. меток, визуально выходящих за пределы доверительного интервала. Было удалено не более одного-двух выбросов из каждой выборки. После усреднения формы выборочная листовая пластина отображалась в виде усредненной симметричной формы – центроида. В результатах Прокрустова анализа (аналога двухфакторного дисперсионного анализа) индивидуальное различие («лист») указывало на различие в форме пластины. Статистически значимый эффект фактора «сторона» указывал на присутствие направленной асимметрии. Взаимодействие обоих факторов указывало на статистическую значимость флуктуирующей асимметрии.
Первый (главный) компонент вариации в матрице «лист×сторона» представлял наибольшую долю дисперсии меток и сравнивались для определения различия в асимметрии. Для нормализации выборок проводилось пермутационное размножение значений координат до 10 000 раз, статистическая значимость определялась на уровне 95 %.
Результаты исследования и их обсуждение
1. Длина и форма листовой пластины
Длина листовых пластин яровой пшеницы была меньше, чем у озимой пшеницы (соответственно 16,7±0,03 и 17,0±0,03 см; df = 667; p << 0,05). Выборки обладали нормальным распределением, так как значения эксцесса и асимметрии находились в пределах {–1÷1}, близкими были как значения коэффициентов вариации, соответственно (3,11±0,03 и 3,23±0,03), так и стандартных отклонений (0,52 и 0,55).
При второй дозе заметно было статистически значимое (p << 0,05) снижение длины листовой пластины яровой пшеницы (16,83±0,05 – 1-я доза и 16,5±0,05 – 2-я доза; рис. 2).
У озимой пшеницы различия в длине пластины не было или оно было слабым (p ≥ 0,05). В контроле длина пластин обоих сортов была заметно больше, чем после внесения второй дозы удобрения (p = 0,001). Авторы связывают это с превышением синтетических процессов увеличения биомассы колоса над фотосинтетической активностью и митотической активностью в листовой пластине.
Величина центроидной фигуры была также меньше у яровой пшеницы (0,076 и 0,080 – озимая; df = 667; p < 0,01). Прямой корреляции или регрессионной зависимости между длиной пластины и величиной центроида (средний квадрат расстояния от координат до геометрического центра) получено не было (p > 0,05).
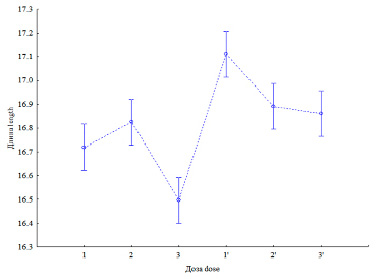
Рис. 2. Влияние двух доз удобрения на длину пластины (95 %-ный доверительный интервал). 1 – яровая контр.; 2 – яровая N60P60K60; 3 – яровая N90P90K90 (p <<0,05) и 1’ – озимая контр. 2’ – озимая N60 + 40 т навоза/га; 3’ – озимая N60 + 40 т навоза/га + N40P40K40 (p ≥ 0,05)
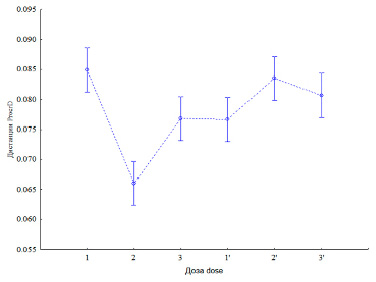
Рис. 3. Влияние двух доз удобрения на величину усредненной фигуры (95 %-ный доверительный интервал). 1 – яровая контр.; 2– яровая N60P60K60; 3 – яровая N90P90K90 (p <<0,05) и: 1’ – озимая контр. 2’ – озимая N60 + 40 т навоза/га; 3’ – озимая N60 + 40 т навоза/га + N40P40K40 (p ≥ 0,05)
Таблица 1
Различие в форме (лист), в направленной асимметрии (сторона) и во флуктуирующей асимметрии (лист × сторона)
|
Источник вариации |
Озимая |
Яровая |
|||||||
|
Effect |
SS |
MS |
df |
F |
SS |
MS |
df |
F |
|
|
Контроль |
лист (1) |
0,017 |
0,00001 |
2700 |
2,09*** |
0,013 |
0,000005 |
2700 |
2,72*** |
|
сторона (2) |
0,000 |
0,00000 |
100 |
0,56ns |
0,0003 |
0,000003 |
100 |
1,85*** |
|
|
(1)×(2) |
0,008 |
0,00000 |
2700 |
2,22*** |
0,005 |
0,000002 |
2700 |
1,63*** |
|
|
ошибка (3) |
0,023 |
0,00000 |
16800 |
0,018 |
0,000001 |
16400 |
|||
|
1-я доза |
(1) |
0,017 |
0,00001 |
2700 |
1,37*** |
0,015 |
0,00001 |
2700 |
1,49*** |
|
(2) |
0,001 |
0,00001 |
100 |
1,88*** |
0,002 |
0,00002 |
100 |
4,34*** |
|
|
(1)×(2) |
0,012 |
0,00000 |
2700 |
2,67*** |
0,010 |
0,00000 |
2700 |
4,04*** |
|
|
(3) |
0,028 |
0,00000 |
16600 |
0,016 |
0,00000 |
16800 |
|||
|
2-я доза |
(1) |
0,014 |
0,00001 |
2700 |
1,32*** |
0,017 |
0,00 |
2 700 |
1,64*** |
|
(2) |
0,000 |
0,00000 |
100 |
0,71ns |
0,001 |
0,00001 |
100 |
1,41* |
|
|
(1)×(2) |
0,011 |
0,00000 |
2700 |
4,82*** |
0,010 |
0,00 |
2 700 |
3,77*** |
|
|
(3) |
0,014 |
0,00000 |
16800 |
0,017 |
0,00000 |
16 800 |
|||
Примечание. SS – сумма квадратов; df – степень свободы; MS – средний квадрат; F – критерий Гудолла; *** – p < 0,0001; * – p < 0,01.
У яровой пшеницы зависимость величины центроида от дозы удобрения была высокой (F = 22,9; р << 0,05), у озимой – практически незначимой (F = 3,8; р = 0,02; рис. 3).
Таким образом, связь между длиной, формой пластины и дозой удобрения была характерна для яровой пшеницы. Для озимой пшеницы серьезной связи между дозой и формой пластины получено не было.
2. Асимметрия листовых пластин в Прокрустовом анализе
Результаты Прокрустова дисперсионного анализа представлены в табл. 1.
Величина ФА (лист × сторона) находилась на одном уровне статистической значимости (p < 0,0001), и была заметна тенденция к повышению во второй дозе удобрения по сравнению с контролем. У озимой формы критерий Гудолла повышался от F = 2,22 до 4,82 и у яровой от F = 1,63 до 3,77.
Направленная асимметрия (сторона) была более характерна для яровой пшеницы, что затрудняло определение величины ФА и стабильности развития.
Индивидуальное различие формы (лист) варьировало от больших значений в контроле (F = 2,09 – озимая и F = 2,72 – яровая) до более низких значений F во второй дозе, соответственно F =1,32 – озимая и F =1,64 – яровая. Авторы относят это явление к равномерному распределению питательных элементов и выравниванию формы пластины в процессе получения важных минералов – азота, фосфора и калия.
У озимой пшеницы сила связи RV между величиной ФА и центроидным размером была незначимой: RV = 0,02 − 0,03 (p > 0,05). Проверка связи ФА – форма у яровой пшеницы показала высокую связь: RV = 0,06 – 0,15 (p < 0,001), то есть отчетливее прослеживалась связь между дозой, формой и ФА, что авторами данной статьи было отнесено к генетическим и морфологическим особенностям яровой пшеницы.
3. Определение различия в асимметрии с помощью морфогеометрических расстояний
Канонический вариационный анализ выполнялся на основе ковариационных матриц координат ХУ меток взаимодействия «лист × сторона». Использовались первые наиболее важные компоненты, занимавшие более 70 %. Между центрами значений в Прокрустовом усредненном пространстве рассчитывались расстояния, указывавшие на их различие (табл. 2).
Статистически значимое различие в асимметрии получено только у озимой пшеницы: 1-я доза – контроль (0,009; p < 0,001) и 2-я доза – контроль (0,006; p < 0,001).
Таблица 2
Прокрустовы расстояния между центрами множеств
|
Озимая |
Яровая |
|||||
|
1 доза |
2 доза |
1 доза |
2 доза |
|||
|
1 доза |
х |
х |
1 доза |
х |
х |
|
|
2 доза |
0,003 (p ≤ 0,05) |
х |
2 доза |
0,003 (p > 0,05) |
х |
|
|
контр. |
0,009 (p < 0.0001) |
0,006 (p < 0.0001) |
контр. |
0,003 (p > 0,05) |
0,002 (p > 0,05) |
|
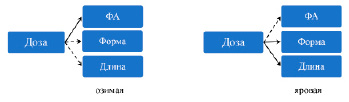
Рис. 4. Связь между дозой и морфогеометрическими характеристиками листовой пластины. Сплошная стрелка показывает значимую связь (р <0,05), пунктирная стрелка – отсутствие или слабую связь (р ≥ 0,05)
Различие в ФА в морфогеометрическом пространстве с помощью канонических переменных между центрами множеств координат было больше выражено у озимой пшеницы.
Таким образом, озимая пшеница показала статистически значимое разделение множеств, т.е. анализ главных компонентов подтверждал влияние дозы удобрения на величину ФА пластин. Различие между контролем и экспериментом в дистанциях Махаланобиса, с учетом эллипсоидного распределения, было у озимой – 3,2, у яровой – 2,0 (p < 0,0001). Пластины озимого сорта различались по асимметричности, как и по результатам Прокрустова анализа (низкие значения НА и высокие ФА), так и в вариационном анализе. В противоположность, листья яровой пшеницы имели меньшую длину, варьировали по форме и обладали высокой направленной асимметрией.
Заключение
Влияние длины на форму не ограничивается лишь одной длиной, а может зависеть и от других неисследованных факторов. Антибатное влияние дозы на длину пластины было получено только у яровой пшеницы. Автор объясняет это коротким вегетативным периодом яровой пшеницы с формированием анатомических структур с широким диапазоном изменчивости, в том числе флуктуационной изменчивости, с высоким стандартным отклонением величины ФА. Таким образом, высокая гетерогенность формы влияла на флуктуирующую асимметрию пластин яровой пшеницы.
В работе, выполненной в Калужской области, было получено увеличение ФА при повышенной дозе минеральных удобрений. В предлагаемой работе результаты анализа главных компонентов подтвердили такую тенденцию.
Отличие в величине ФА озимой пшеницы, полученной при разных дозах удобрения, означало влияние высокоинтенсивного органоминерального удобрения на асимметричность листовых пластин. Морфологическое различие пластин двух форм пшеницы отражало генетическое содержание асимметрии. На это указывало и различное соотношение ФА/НА у исследованных сортов. Связь длина – форма – ФА сильнее проявлялась у яровой пшеницы (рис. 4).
Генетические особенности и разные сроки посева не позволяли четко сопоставлять обе формы пшеницы по стабильности развития. Высокоинтенсивная доза могла способствовать неравномерному росту клеток в пластинах яровой пшеницы, что объясняет высокое присутствие направленной асимметрии, включая и контрольную выборку.
Ранее на примере древесных растений продемонстрировано, что пластическая и флуктуационная изменчивость находились в обратной связи. Напротив, изучение мерных признаков пшеницы не показало такой зависимости. В сравнении с озимой пшеницей пластическая изменчивость представляется скорее свойством яровой пшеницы, которое сопряжено с флуктуационной изменчивостью. Последующее изучение будет направлено на определение асимметрии при воздействии других доз удобрения и влияния климатических факторов.